
Introduction: Why the Access Method Is as Important as the Target
The previous articles in this series established the neuroanatomical rationale for the otic ganglion as the primary juncture point of the cervical crush pattern, mapped the convergence of mechanical, vascular, autonomic, and neurological forces that produce the hidden pulse, and connected the classical TCM description of Yin-Yang separation to its precise anatomical substrate.
This article addresses the clinical question that follows: given that these structures are the target, how do we access them without triggering the very threat detection systems whose dysregulation we are attempting to address?
The answer to that question is the foundation of the manual approach described here. In this population — characterized by chronic dorsal vagal dominance, kindled PAG activation, retained primitive reflexes, and a nervous system primed for shutdown or explosion in response to threat input in the craniofacial territory — the access method is not secondary to the anatomical target. It is co-equal with it. An anatomically precise intervention delivered through the wrong sensory modality will produce the opposite of its intended effect before any therapeutic mechanism has had the opportunity to operate.
The face and skull acupuncture points sit directly over the parasympathetic ganglia, trigeminal emergence points, cranial nerve exit foramina, and autonomic convergence points mapped in the previous article. The classical system mapped the clinical effects through centuries of observation. The neuroanatomy explains the mechanism. What both traditions share is the identification of where to work. What the current synthesis adds is a precise understanding of how the access method must be chosen for this specific population — and why the choice between touch, needles, and electrical stimulation is not a matter of practitioner preference but of neurological safety.
Part One: The Threat Detection Filter — Why Access Method Determines Outcome
The Craniofacial Territory as Highest-Density Threat Zone
The face, skull, and upper cervical region above T3 is the highest concentration zone of:
Primitive reflex receptive fields. The perioral, rooting, Moro, tonic labyrinthine, and tonic neck reflexes all have their primary sensory receptive fields in the face, head, and upper cervical territory. Any sharp or unexpected sensory input in this territory risks activating retained primitive reflexes involuntarily — not as a therapeutic effect but as an uncontrolled defensive activation that drives the system further into the freeze or flight state it already occupies.
Trigeminal nociceptive territory. The trigeminal system innervates the entire face and head with one of the highest receptor densities in the body. Sharp painful input in trigeminal territory produces immediate central sensitization in the trigeminal nucleus — which spans the entire brainstem and connects directly to the vagal nuclei and autonomic regulatory centers. Central sensitization in the trigeminal nucleus is the opposite of the autonomic downregulation the intervention is attempting to produce.
TCR trigger zones. The periorbital, nasal, perioral, and oropharyngeal territories where the TCR is most readily activated are all within this zone. Nociceptive input in these territories risks triggering the TCR pathway and producing cardiac autonomic effects — bradycardia, blood pressure instability — that are unpredictable and potentially destabilizing in a population whose cardiac autonomic regulation is already compromised.
Freeze and startle reflex territory. The neck is the primary territory of the tonic labyrinthine and ATNR reflexes. Sharp painful input to the cervical muscles triggers the protective withdrawal reflex — which in a nervous system with retained primitive reflexes activates the full reflex rather than a graduated response.
The Sensory Hierarchy and the PAG Threshold
The kindled PAG in this population — described in detail in Article One — has a lowered activation threshold for the freeze response. Any sensory input that reaches the threat-coded level activates the PAG before cortical processing can participate. The therapeutic intention is irrelevant. The nervous system responds to the sensory quality of the input, not to its intended purpose.
This creates a specific clinical requirement: every intervention in the craniofacial territory above T3 must be evaluated first through the threat detection lens. The question is not whether the intervention produces the desired effect in a normally regulated nervous system. The question is what the intervention does to the threat detection system of this specific nervous system before any therapeutic effect has the opportunity to occur.
The sensory hierarchy in this territory, from lowest to highest threat activation potential, runs approximately as follows:
- Sustained gentle non-moving contact — lowest threat activation, activates C-tactile afferents, Ruffini endings, and slow-adapting mechanoreceptors associated with parasympathetic activation.
- Slow gentle moving touch — activates C-tactile afferents specifically, produces oxytocin release and direct insular cortex activation, remains below nociceptive threshold.
- Firm sustained pressure — crosses into pressure receptor activation, still below nociceptive threshold with adequate approach, but requires the nervous system to have sufficient regulatory capacity to tolerate pressure without reading it as threat.
- Sharp or unexpected contact — activates A-delta fibers and nociceptors, involuntarily triggers withdrawal reflexes, activates the primitive reflex circuits, reaches the PAG at or above its lowered threshold.
- Penetrating contact (needling) — activates all nociceptive pathways simultaneously, unavoidably triggers the primitive reflex circuits in the reflexogenic zones of the face and scalp, activates the TCR through nociceptive input in periorbital and nasal territory, central sensitization in trigeminal nucleus.
- Electrical stimulation — all of the above plus direct neural membrane depolarization, potential cortical spreading depression in brain trauma tissue, kindled PAG activation through electrical nociception, voltage-gated calcium channel dysregulation in neural tissue.
The clinical principle that follows is absolute for this population: above T3, particularly from the clavicle upward, hands and non-intrusive touch; needles are ineffective and potentially harmful; electrical stimulation is contraindicated.
Part Two: Why Needles Are Ineffective Above T3 in This Population
The Needle as Unavoidable Threat Trigger
A needle is by definition a sharp, unexpected, penetrating stimulus. Even in a cooperative, willing adult, needle penetration activates A-delta pain fibers, C fiber nociceptors, the withdrawal reflex arc (involuntary), the orienting response (involuntary), potentially the TCR for periorbital and nasal territory needling, and the primitive reflex circuits for any stimulation in the reflexogenic zones of the face and scalp.
In a normally regulated nervous system these activations are brief and manageable. The cortical regulatory systems modulate the response. The practitioner observes a brief needle sensation followed by the deqi response and tissue settling.
In this population the picture is categorically different:
- The cortical regulatory systems are chronically occupied suppressing the limbic system and have minimal spare regulatory capacity for managing acute nociceptive input.
- The PAG is kindled and has a lowered threshold for freeze activation — the nociceptive input from the needle in craniofacial territory reaches the PAG activation threshold and triggers the freeze cascade before any cortical modulation can participate.
- The primitive reflexes are retained and will fire involuntarily to the needle stimulus — not as a therapeutic reflex but as an uncontrolled defensive activation.
- The trigeminal central sensitization means the needle input produces amplified central processing rather than normal proportionate response — the central sensitization generated is the opposite of therapeutic.
The threat template means the needle penetration in the face and skull is processed through the threat detection system before any conscious cognitive appraisal can participate.
The consequence is not simply that the needle fails to produce the intended therapeutic effect. It is that the needle actively drives the system deeper into the defensive state it already occupies — increasing sympathetic or dorsal vagal activation, reinforcing the cervical crush pattern through defensive muscular bracing, and potentially triggering the TCR with unpredictable cardiac consequences in a compromised autonomic system.
Unskilled or painful needling in this territory will trigger primitive reflexes whether the practitioner intends it or not. Pain is a reflex trigger. This is involuntary and the practitioner’s therapeutic intention does not override the neurological response. This is not a soft clinical preference. It is a neurological fact with direct patient safety implications.
Part Three: Why Electrical Stimulation Is Contraindicated Above T3 in Brain Trauma and Developmental Trauma Populations
The Mechanism of Harm
Electrical stimulation on the scalp for brain trauma is not merely suboptimal in this population. There are specific mechanistic reasons why it is likely actively harmful and why the field’s continued use of it represents a significant clinical safety issue.
Direct TCR activation. Electrical stimulation on the scalp activates the trigeminal sensory system through the cutaneous branches covering the scalp — supraorbital and supratrochlear branches of V1, zygomaticotemporal branch of V2, auriculotemporal branch of V3. Through the TCR pathway, this produces direct vagal motor nucleus activation — potentially generating bradycardia, arrhythmia, and in a sensitized system the full TCR cascade. In a brain trauma patient whose brainstem autonomic regulation is already compromised by the injury, TCR activation from scalp estim is not a trivial peripheral event. It is driving directly into the most vulnerable regulatory center.
Kindled PAG activation. Electrical stimulation activates nociceptors even at sub-threshold intensities. In the brain trauma or developmental trauma population with kindled PAG, nociceptive input from scalp estim activates the PAG through the trigeminal-PAG pathway at levels far below what would produce visible pain behavior. The PAG activation produces the freeze-shutdown cascade — which is neurologically the opposite of what brain trauma rehabilitation requires.
Repeated primitive reflex activation. Electrical stimulation in cranial territory activates the same primitive reflex circuits that needle stimulation activates — with the additional variable of electrical current spreading through tissue and potentially activating reflex circuits at a distance from the electrode placement. Retained primitive reflexes in the brain trauma population — which are extremely common because brain trauma disrupts the cortical inhibition of primitive reflexes — fire involuntarily to the electrical stimulus. Repeated electrical activation of retained primitive reflexes does not integrate them. It re-triggers them — potentially reinforcing the retained reflex architecture rather than resolving it.
Cortical spreading depression. Electrical stimulation on the scalp can trigger cortical spreading depression — a wave of neuronal depolarization followed by suppression propagating across the cortex. In normal tissue this is transient. In brain trauma tissue with compromised ionic homeostasis and disrupted blood-brain barrier integrity, cortical spreading depression can be more extensive, more prolonged, and more damaging. It is a known mechanism of secondary injury in traumatic brain injury. Scalp estim applies electrical current to tissue that may be primed to generate it.
Blood-brain barrier compromise. Brain trauma disrupts the blood-brain barrier. Electrical stimulation drives ionic current through scalp and skull into brain tissue. In compromised blood-brain barrier the current is not filtered and buffered normally. Ionic shifts in the perilesional tissue — already metabolically compromised — may produce additional dysregulation in precisely the tissue most vulnerable to it.
Inflammatory amplification. Electrical stimulation produces local tissue inflammation through mast cell degranulation and inflammatory mediator release at electrode sites. In scalp overlying brain trauma territory this local inflammatory response adds to the already elevated neuroinflammatory load. Inflammatory mediators enter the lymphatic and vascular drainage of the scalp and contribute to the systemic inflammatory picture.
Why It Continues to Be Used
The persistence of scalp estim in brain trauma rehabilitation despite these contraindications reflects several converging problems: research supporting scalp estim was largely conducted on neurologically intact populations — stroke patients with intact blood-brain barriers and normally regulated nervous systems — without adequately accounting for the mechanistic differences between stroke and traumatic brain injury; the visible muscle twitching and measurable cortical activation creates the appearance of therapeutic effect even when the underlying mechanism may be counterproductive; practitioners using it are largely not trained in the autonomic neuroscience that would allow recognition of the TCR activation, PAG kindling, and primitive reflex triggering occurring in this population; and adverse effects are not systematically tracked in ways that would reveal the pattern.
The clinical principle is clear: no electrical stimulation in the craniofacial territory for brain trauma patients, developmental trauma patients, patients with retained primitive reflexes, or any patient presenting with the hidden pulse pattern and chronic dorsal vagal dominance described in this series.
Part Four: The Mechanoreceptor and Piezoelectric Basis for Non-Intrusive Touch
Why Sustained Gentle Contact Works at the Tissue Level
Non-intrusive sustained touch produces its therapeutic effects through a specific sequence of tissue responses that are distinct from the effects of needle stimulation or electrical stimulation. Understanding this sequence provides the practitioner with the mechanistic rationale for the specific quality, duration, and pressure of contact that the technique requires.
Zero to two seconds — elastic tissue response. Initial mechanical deformation of fascial tissue. The viscoelastic properties of fascia mean it initially resists deformation — the elastic component responds first. The tissue pushes back against the contact. No therapeutic tissue change is occurring yet. This is the practitioner learning to simply rest their contact against the tissue’s own resistance without forcing through it.
Two to five seconds — Ruffini ending activation. Sustained pressure begins engaging the Ruffini mechanoreceptors — slowly adapting mechanoreceptors in the dermis and deep fascia that respond specifically to sustained lateral stretch and compression. Ruffini ending activation produces a documented reduction in sympathetic tone in the surrounding tissue (Schleip, 2003). They are mechanically-activated parasympathetic switches embedded in the fascial tissue — activated not by movement or sharp stimulus but by sustained gentle deformation. This is the tissue beginning to respond.
At approximately five seconds — the piezoelectric threshold. Collagen fibers under sustained mechanical deformation generate piezoelectric currents — electrical signals produced by mechanical stress on crystalline collagen structures. Fascia has significant piezoelectric properties because of its collagen content and its liquid crystal molecular organization (Langevin, 2006). The piezoelectric current produced by sustained compression propagates through the fascial continuity — in this case through the sinew channel architecture of the cervical and craniofacial region. The practitioner may feel this as a subtle warmth, a pulsation, or a softening beginning in the tissue under their fingers — the beginning of what clinically presents as the tissue melt.
The tissue melt — fascial creep response. The point at which the ground substance of the connective tissue shifts from its more gel-like state toward a more sol-like state under sustained mechanical input. This is a real phase transition in the extracellular matrix mediated by mechanotransduction through integrins — the transmembrane proteins connecting extracellular matrix to the intracellular cytoskeleton — triggering intracellular signaling cascades that alter cellular contractility in fibroblasts and smooth muscle; hyaluronic acid viscosity changes with mechanical input and temperature; piezoelectric current propagation altering the local tissue electrical environment; and mast cell degranulation releasing local mediators that alter tissue viscosity.
The tissue melt is not a subjective perception. It is an objectively occurring change in the physical state of the connective tissue ground substance. The practitioner feels it because the resistance under their fingers literally changes — the tissue transitions from pushing back against the contact to moving with it.
C-Tactile Afferents and Featherweight Touch
For the most superficial and sensitive zones — particularly the clavicular triangle and the supraclavicular fossa — featherweight touch activates a specific class of sensory fiber not activated by firmer contact: C-tactile afferents. These unmyelinated sensory fibers respond specifically to gentle, slow, moving touch. They project to the insular cortex and produce direct parasympathetic activation and oxytocin release (McGlone et al., 2014). They have a specific activation threshold that is below the threshold of pressure receptors and far below the threshold of pain receptors.
Featherweight touch is not a softer version of regular touch. It is activating a categorically different sensory pathway that produces categorically different autonomic effects. For the territory of ST 12-13 in the clavicular triangle — where the brachial plexus is superficial and any pressure that creates a nociceptive response would trigger upper extremity withdrawal reflexes — featherweight touch is the only contact quality that accesses the therapeutic pathway without simultaneously triggering the defensive pathway.
Part Five: The Four Parasympathetic Ganglia of the Head — Surface Access Points
Before mapping the specific hand positions, the four parasympathetic ganglia of the head are identified with their anatomical locations and acupuncture point references, as these constitute the primary therapeutic targets of the craniofacial territory.
The Ciliary Ganglion
Anatomical location: Sits in the posterior orbit approximately one centimeter anterior to the optic foramen, lateral to the optic nerve, between the optic nerve and the lateral rectus muscle. Receives preganglionic parasympathetic fibers from the Edinger-Westphal nucleus via the inferior division of CN III. Sends postganglionic fibers to the sphincter pupillae and ciliary muscle. Also carries the nasociliary branch of the trigeminal nerve — the sensory fiber source for the oculocardiac reflex.
Surface access points: BL 1 — Jingming — at the medial canthus, closest surface point to the posterior orbit, accessing the nasociliary nerve territory that activates the TCR. BL 2 — Zanzhu — at the medial end of the eyebrow, supraorbital notch, accessing the supraorbital branch of V1. ST 1 — Chengqi — below the pupil on the infraorbital rim. GB 1 — Tongziliao — at the lateral canthus. The complete orbital ring — BL 1, BL 2, ST 1, ST 2, GB 1, TB 23, Yuyao — maps onto the trigeminal sensory territory feeding into the ciliary ganglion and the TCR pathway.
The Pterygopalatine Ganglion
Anatomical location: Sits in the pterygopalatine fossa, the small pyramidal space just posterior to the maxilla, inferior to the orbital apex, lateral to the perpendicular plate of the palatine bone, and directly adjacent to the sphenoid body. Receives preganglionic parasympathetic fibers from the facial nerve via the greater petrosal nerve. Sends postganglionic fibers to the lacrimal gland, nasal mucosa, palatine glands, and pharyngeal mucosa.
Surface access points: ST 7 — Xiaguan — is the most direct surface approach, sitting in the depression anterior to the condylar process and inferior to the zygomatic arch, providing access to the infratemporal fossa region where the pterygopalatine fossa is located at depth. LI 20 and Bitong access the nasal mucosal territory this ganglion innervates. The nasal approach relates to the territory through which sphenopalatine ganglion blocks are performed in interventional pain medicine.
The Submandibular Ganglion
Anatomical location: Suspended from the lingual nerve — a branch of V3 — sitting just superior to the submandibular salivary gland in the floor of the mouth. Receives preganglionic parasympathetic fibers from the facial nerve via the chorda tympani joining the lingual nerve. Sends postganglionic fibers to the submandibular and sublingual salivary glands.
Surface access points: The submandibular triangle between CV 23 — Lianquan — ST 5 — Daying — and the midline corresponds to the anatomical location of the ganglion and its glandular targets. ST 6 — Jiache — at the masseter belly, and the anterior border of the masseter in the ST 5 region, access the mandibular and submandibular territory.
The Otic Ganglion
Anatomical location: Sits just inferior to the foramen ovale, medial to the mandibular nerve, immediately below the base of the skull in the infratemporal fossa, at approximately the level of the neck of the mandibular condyle. Receives preganglionic parasympathetic fibers from CN IX via the lesser petrosal nerve. Sends postganglionic fibers to the parotid gland via the auriculotemporal nerve. Sits at the anatomical junction of the parasympathetic and sympathetic inputs to the cranial territory — the primary juncture point described in Article Two.
Surface access points: The triangle formed by ST 7 — Xiaguan — GB 2 — Tinghui — and TB 21 — Ermen — approximates the infratemporal fossa territory where the otic ganglion resides. SJ 17 — Yifeng — posterior to the earlobe in the depression between mastoid and mandible, sits slightly posterior to the otic ganglion itself but accesses the parotid territory, the posterior infratemporal fossa, and the stylomastoid foramen where the facial nerve exits — connecting to the parasympathetic system through the facial nerve’s preganglionic branches.
Part Six: The First Hand Position — Skull Base to C4
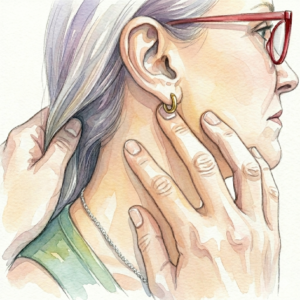 The Clinical Context The first hand position addresses the cervical crush pattern from the skull base down to C4, opening the superior aspect of the compressed cylinder, contacting three of the four cranial parasympathetic ganglia, decompressing the carotid sheath superiorly, and initiating the piezoelectric cascade through the craniofacial sinew channel architecture. It is typically applied to one side at a time — left hand working the left jaw-neck complex, right hand working the right.
The Clinical Context The first hand position addresses the cervical crush pattern from the skull base down to C4, opening the superior aspect of the compressed cylinder, contacting three of the four cranial parasympathetic ganglia, decompressing the carotid sheath superiorly, and initiating the piezoelectric cascade through the craniofacial sinew channel architecture. It is typically applied to one side at a time — left hand working the left jaw-neck complex, right hand working the right.
The five contact points of the first hand position and their precise neuroanatomical territory are mapped below.
Middle Finger — SJ 17 (Yifeng)
Surface location: Posterior to the earlobe in the depression between the mastoid process and the posterior border of the mandible.
Anatomical territory contacted:
The stylomastoid foramen territory — the point of facial nerve exit from the skull, the facial nerve carrying preganglionic parasympathetic fibers to both the pterygopalatine and submandibular ganglia through its branches.
The posterior belly of digastric and stylohyoid — key components of the hyoid chain whose chronic tension contributes to the anterior cervical crush pattern.
The parotid gland’s posterior border — the parotid being the postganglionic target of the otic ganglion through the auriculotemporal nerve.
The otic ganglion territory at depth — the otic ganglion sitting approximately one to two centimeters anterior, superior, and medial to this contact point at the level of foramen ovale inside the skull base.
The great auricular nerve — C2-C3 — the superficial cervical plexus branch covering the parotid and mastoid region, connecting to the trigeminocervical complex.
The jugular foramen territory at depth — CN IX, X, XI and internal jugular vein exiting the skull immediately adjacent.
Clinical significance: SJ 17 is the most posterior access point to the otic ganglion territory and the jugular foramen complex. It contacts the primary autonomic juncture point described in Article Two from its most accessible posterior approach, and provides access to the facial nerve exit point whose preganglionic branches supply two of the four parasympathetic ganglia.
Ring Finger — ST 6 (Jiache)
Surface location: At the belly of the masseter, at the angle of the mandible.
Anatomical territory contacted:
The masseter muscle belly — typically carrying the highest chronic tension of any craniofacial muscle in freeze-dominant and threat-hypervigilant presentations. The masseter is one of the primary expressions of the V3 motor lock described in the published Polyvagal Acupuncture® article.
The masseter nerve — a branch of V3 — carrying motor supply to the masseter and proprioceptive feedback into the trigeminal system. This is a direct entry point into the V3 motor lock circuit.
The parotid duct emerging from the anterior parotid in this territory.
The pterygomandibular space at depth — where the inferior alveolar nerve and lingual nerve run — deep V3 territory connecting to both the TCR pathway and the submandibular ganglion.
The facial artery crossing the mandible just anterior to the masseter.
Clinical significance: ST 6 in combination with SJ 17 brackets the parotid gland and the masseter, simultaneously contacting the otic ganglion territory posteriorly (SJ 17) and the V3 motor-sensory territory anteriorly (ST 6). These two fingers together are holding the jaw’s autonomic and myofascial architecture between them — the posterior parasympathetic ganglion territory and the anterior trigeminal motor lock.
Pinky — Submandibular Triangle
Surface location: Moving inferior to the mandibular angle into the submandibular triangle, toward the region between CV 23 and ST 5.
Anatomical territory contacted:
The submandibular ganglion territory — the third of the four parasympathetic ganglia, accessed from its most superficial external approach.
The submandibular salivary gland.
The facial artery as it curves around the mandible.
The mylohyoid — the primary floor-of-mouth muscle whose tension contributes to the anterior cervical floor compression.
The anterior belly of digastric.
The lingual nerve carrying both V3 sensory fibers and chorda tympani parasympathetic fibers to the submandibular ganglion.
The hypoglossal nerve — CN XII — running just deep to the digastric tendon.
Clinical significance: Adding the pinky contact means three of the four cranial parasympathetic ganglia — otic (via SJ 17), pterygopalatine (via ST 7 territory accessed through ST 6 depth), and submandibular (via the submandibular triangle) — are being simultaneously contacted in their most superficially accessible territory. The three-ganglion contact creates a field of sustained gentle parasympathetic input that is qualitatively different from single-point contact — the nervous system is receiving simultaneous invitation to parasympathetic activation from multiple ganglia simultaneously.
Index Finger — Between GB 12 and GB 20
Surface location: On the posterior occiput between GB 12 (Wangu — posterior and inferior to the mastoid process) and GB 20 (Fengchi — in the depression between sternocleidomastoid and trapezius at the occiput).
Anatomical territory contacted:
The suboccipital region — the attachment territory of the suboccipital muscles (rectus capitis posterior major and minor, obliquus capitis superior and inferior). These are the posterior component of the cervical crush pattern — their release through sustained contact is the primary posterior element of the first hand position.
The posterior atlanto-occipital membrane.
The greater occipital nerve — C2 dorsal ramus — which has documented connections to the trigeminal system through the trigeminocervical complex in the upper cervical spinal cord. The trigeminocervical complex convergence means that sustained contact on the greater occipital nerve territory produces effects that propagate into the trigeminal sensory nucleus and thence to the TCR pathway and the nucleus tractus solitarius.
The occipital condyle region at depth — the condyle sitting directly superior to the jugular foramen. Release of the posterior atlanto-occipital compression through this contact directly reduces the posterior compressive force on the jugular foramen from above.
The vertebral artery territory — the posterior atlanto-axial joints and the suboccipital space containing the vertebral arteries as they curve medially before entering the foramen magnum. Release of suboccipital compression through this contact improves vertebral artery flow and posterior circulation perfusion to the brainstem autonomic nuclei.
Clinical significance: The index finger on the posterior occiput is the posterior anchor of the entire first hand position. It contacts the posterior component of the cervical crush at the skull base — the mirror image of the anterior crush forces accessed by the other fingers. Together, the SJ 17-ST 6-submandibular-GB 12/20 contact pattern is spanning the anterior and posterior components of the cervical crush at the skull base level simultaneously.
Thumb — C4 (SI 16 / LI 17 Territory, Posterior SCM Border)
Surface location: On the posterior border of the sternocleidomastoid at approximately the C4 level, in the region of SI 16 — Tianchuang — with the Sanjiao empirical point for the carotid sinus territory in the immediate vicinity.
Anatomical territory contacted:
The carotid bifurcation. The common carotid artery bifurcates into internal and external carotid at approximately C3-C4. The carotid sinus — the baroreceptor at the bifurcation regulating blood pressure and heart rate through CN IX — sits in this territory. Gentle sustained pressure on the carotid sinus produces direct vagal activation — it is used clinically to terminate certain supraventricular tachycardias. The thumb at C4 is on the primary peripheral input to the cardiac vagal reflex arc.
The carotid body. The chemoreceptor sensing blood oxygen, CO2, and pH, sitting at the carotid bifurcation. In the chronic hypoxia picture described in Article Two, the carotid body is detecting and reporting reduced oxygen tension to the brainstem through CN IX and the nucleus tractus solitarius. The thumb at C4 is on the peripheral sensor reporting the very hypoxia the technique is attempting to resolve.
The vagal cardiac branches. The vagus nerve gives off its superior and middle cervical cardiac branches — the parasympathetic decelerator fibers to the heart — at this level in the carotid sheath.
The cervical sympathetic cardiac nerves. The cervical sympathetic trunk gives off its cardiac accelerator branches at this level — the sympathetic counterpart to the vagal decelerator fibers. Both the parasympathetic cardiac output and the sympathetic cardiac output are running through the carotid sheath territory at C4 under the thumb simultaneously.
The phrenic nerve. Formed at C4 on the anterior surface of scalenus anterior. Carries motor supply to the diaphragm and sensory fibers from the pericardium. As the scalene tension releases under the thumb contact, phrenic nerve decompression allows diaphragmatic function to normalize — improving intrathoracic pressure dynamics, venous return, and the respiratory sinus arrhythmia component of HRV. The pericardial sensory feedback that contributes to cardiac regulatory input begins to restore.
The anterior scalene. One of the primary muscles of the thoracic outlet compression and the cervical crush pattern. Its release at the C4 level begins the opening of the thoracic inlet that the second hand position completes inferiorly.
Clinical significance: The thumb at C4 is the peripheral cardiac autonomic hub of the entire first hand position. It simultaneously contacts the primary baroreceptor input to cardiac vagal regulation, the afferent cardiac sensory pathway through the phrenic nerve, the efferent cardiac parasympathetic output through vagal cardiac branches, the efferent cardiac sympathetic output through cervical sympathetic cardiac nerves, and the oxygen chemoreceptor detecting the chronic hypoxia. The entire peripheral cardiac autonomic regulatory circuit — input and output, sympathetic and parasympathetic — is concentrated at C4 under the thumb.
The Complete First Hand Position
With all five contacts simultaneously established the hand is spanning:
Posteriorly — suboccipital release point, greater occipital nerve, trigeminocervical complex, vertebral artery territory (index finger)
Laterally at skull base — otic ganglion territory, jugular foramen complex, facial nerve exit, posterior carotid sheath (middle finger at SJ 17)
Anterolaterally at jaw — V3 motor-sensory junction, masseter proprioception, parotid territory, TCR sensory input (ring finger at ST 6)
Anteroinferiorly at jaw — submandibular ganglion, floor of mouth, lingual nerve, chorda tympani (pinky in submandibular triangle)
Laterally at C4 — carotid sinus, carotid body, vagal cardiac branches, sympathetic cardiac nerves, phrenic nerve, anterior scalene (thumb at C4)
Three of four cranial parasympathetic ganglia, the primary peripheral cardiac autonomic circuit, the TCR sensory territory, the trigeminal system at multiple entry points, the descending vagus in the carotid sheath, the cervical sympathetic trunk, and the vertebral artery territory are all held simultaneously in sustained non-intrusive contact.
Part Seven: The Second Hand Position — Clavicle to First Intercostal Space
The Clinical Rationale for the Second Hand
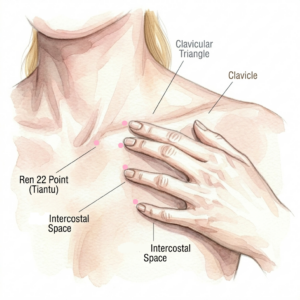 The first hand position opens the superior aspect of the compressed cervical cylinder — addressing the skull base juncture, the carotid sheath compression, and the cervical crush from foramen ovale down to C4. Without a corresponding inferior contact, the release initiated above has no exit pathway. The second hand position provides the inferior anchor — opening the thoracic inlet from above and creating the pressure differential that allows the cascade to flow in its anatomically natural direction: caudally from the cranial vault through the cervical space and into the thoracic cavity.
The first hand position opens the superior aspect of the compressed cervical cylinder — addressing the skull base juncture, the carotid sheath compression, and the cervical crush from foramen ovale down to C4. Without a corresponding inferior contact, the release initiated above has no exit pathway. The second hand position provides the inferior anchor — opening the thoracic inlet from above and creating the pressure differential that allows the cascade to flow in its anatomically natural direction: caudally from the cranial vault through the cervical space and into the thoracic cavity.
The two hands together are spanning the entire length of the compressed cylinder simultaneously — from the otic ganglion juncture at the skull base superiorly to the thoracic inlet inferiorly. This is the anatomical basis for the descending cascade described in the clinical observation that opens the territory out of the head and into the chest.
Middle Finger — Along the Clavicle (Large Intestine Divergent Channel Territory)
Surface location: Laid along the clavicle, corresponding to the territory identified by Royston Lowe as the Large Intestine divergent channel.
Anatomical territory contacted:
The clavicle is the only bony bridge between the axial and appendicular skeleton — the transition point through which everything passing from the cervical region into the thorax and upper extremity must cross. The clavicular territory carries:
The subclavian artery and vein in the subclavian groove on the inferior clavicular surface.
The brachial plexus trunks passing posterior to the clavicle through the costoclavicular space.
The thoracic duct on the left — emptying at the left subclavian-jugular junction directly posterior to the medial clavicle. The right lymphatic duct on the right at the same location. These are the primary lymphatic drainage junctions where all lymph from the head and neck enters the venous circulation — impaired by the same compression that impairs jugular venous return in the cervical crush pattern.
The clavicular head of SCM attaching to the medial clavicle — the inferior attachment of the primary anterior cervical crush muscle.
The fascial sleeve transition between the deep cervical fascia and the thoracic inlet fascia — the boundary between the cervical and thoracic compartments.
Clinical significance: Royston Lowe’s identification of the Large Intestine divergent channel along the clavicle is anatomically precise. The clavicular territory marks the fascial transition boundary between the cervical and thoracic compartments — the exact barrier the technique is opening to allow the cascade to complete. The middle finger along the clavicle is holding this boundary open as the release descends from above, and simultaneously decompressing the lymphatic drainage junction that the cervical compression has been impairing.
Ring Finger — First Intercostal Space
Surface location: In the first intercostal space between the first and second ribs.
Anatomical territory contacted:
The first intercostal nerve carrying sympathetic preganglionic fibers to the stellate ganglion territory.
The internal thoracic artery and vein running lateral to the sternum.
The endothoracic fascia lining the inner thoracic wall.
The apex of the lung — the pleural cupola extending above the first rib into the root of the neck.
The stellate ganglion territory. The stellate ganglion — the fusion of the inferior cervical and first thoracic sympathetic ganglia — sits at C7-T1 anterior to the first rib’s neck and is the primary sympathetic ganglion supplying the heart, upper extremity, and head and neck. Stellate ganglion block is used clinically for treating PTSD, cardiac arrhythmias, and chronic pain, with documented effects on cardiac sympathetic tone. The first intercostal space is the most superficially accessible territory adjacent to the stellate ganglion.
Clinical significance: The ring finger in the first intercostal space is modulating the stellate ganglion territory — the primary cardiac sympathetic accelerator — while the first hand is simultaneously increasing vagal cardiac parasympathetic tone through the carotid sinus and vagal cardiac branches at C4. The two hands together are modulating both arms of the cardiac autonomic balance — parasympathetic increase superiorly, sympathetic modulation inferiorly — simultaneously and through non-intrusive contact.
Ring Finger Stroke / Featherweight Touch — ST 12-13 First Intercostal
Surface location: Gentle featherweight stroking or sustained touch in the supraclavicular fossa, ST 12 — Quepen — in the center of the supraclavicular fossa, through to ST 13 — Qihu — on the inferior clavicular border at the midclavicular line.
Anatomical territory contacted:
The brachial plexus as it emerges between anterior and middle scalene and passes posterior to the clavicle — the trunks palpable in the supraclavicular fossa.
The phrenic nerve at C4 level in the anterior scalene territory before it descends into the thorax.
The omohyoid muscle crossing the posterior triangle — connecting the hyoid bone to the scapula through a fascial sling that spans the entire cervical-thoracic transition.
The scalene fat pad containing the brachial plexus and vascular structures.
The lymphatic trunks converging toward the thoracic duct junction.
The C-tactile afferent-rich skin of the supraclavicular territory — this is the territory where featherweight touch producing oxytocin release and direct insular cortex activation is most important. Any firmer contact risking brachial plexus nociception would activate upper extremity withdrawal reflexes — the Moro reflex’s upper extremity component — precisely what the technique is designed to avoid.
Clinical significance: The featherweight quality of the ST 12-13 contact is non-negotiable here. The brachial plexus is superficial and palpable. The C-tactile afferents require featherweight touch to activate their specific parasympathetic and oxytocin-releasing pathway. This contact is activating the body’s own social touch system — the pathway through which safe relational contact produces direct autonomic regulation — in the territory most directly overlying the cervical-thoracic transition being opened by the surrounding contacts.
Pinky — ST 14 (Kufang)
Surface location: In the first intercostal space at the midclavicular line.
Anatomical territory contacted:
The clavipectoral fascia — the deep fascia connecting the clavicle to the thoracic wall and enclosing the subclavius and pectoralis minor.
The pectoralis minor — one of the primary muscles of the forward head and rounded shoulder freeze posture, chronically shortened and anteriorly rotating the shoulder girdle. Its chronic shortening contributes to thoracic outlet compression, compresses the neurovascular bundle in the costoclavicular space, and restricts upper rib expansion limiting thoracic breathing depth.
The long thoracic nerve — C5-C6-C7 — innervating serratus anterior, the primary scapular protractor in the freeze shoulder posture.
Clinical significance: The pinky at ST 14 contacts the pectoralis minor territory and clavipectoral fascia — the myofascial anchor of the freeze shoulder posture and a contributor to the thoracic outlet compression. As the tissue melt propagates from the clavicular and first intercostal contacts, the pectoralis minor territory receives the inferior edge of the release wave, beginning the opening of the anterior thoracic freeze armor below the clavicle.
Thumb — Sideways Shear at REN 22, Sternal SCM Attachment
Surface location: At the suprasternal notch, REN 22 — Tiantu — with a sideways shear direction on the sternal head of SCM.
Anatomical territory contacted:
The sternal head of SCM attaching to the anterior surface of the manubrium lateral to REN 22. This is the inferior anchor of SCM — the primary anterior cervical crush muscle whose superior attachment is at the mastoid process, in the territory of the middle finger at SJ 17 in the first hand position.
The two-hand spanning of SCM: With the first hand’s middle finger at SJ 17 contacting the mastoid attachment of SCM superiorly and the second hand’s thumb at REN 22 contacting the sternal attachment inferiorly, the complete length of SCM is spanned between the two hands simultaneously. This is the primary mechanical mechanism of the descending cascade.
The pretracheal fascia anterior to the trachea.
The anterior jugular veins and their connecting arcade.
The thyroid cartilage and its fascial attachments.
The sideways shear specifically: Fascia responds differently to shear forces than to direct compression. Shear loading produces lateral displacement of collagen fibers relative to each other — the primary mechanism for releasing fascial adhesions between tissue layers. It activates Golgi tendon organ-like mechanoreceptors at the musculotendinous junction — producing autogenic inhibition of the muscle, a reflex relaxation of SCM through its own proprioceptive feedback. And it creates piezoelectric currents oriented differently from those produced by direct compression — providing a directional bioelectrical stimulus through the SCM fascial envelope and the connected cervical fascial sleeve.
The sideways shear at the sternal SCM attachment with simultaneous hold at the mastoid attachment produces longitudinal shear through the entire SCM — from sternum to mastoid — that autogenically inhibits the entire muscle, mechanically decompresses the carotid sheath through the lateral displacement produced by the shear, and creates longitudinal piezoelectric current propagation through the SCM fascial envelope connecting to the entire cervical fascial sleeve.
Part Eight: The Descending Cascade — Mechanism and Clinical Observation
What the Cascade Is
The descending movement observed in clinical practice — beginning at the otic ganglion territory and cascading down through the clavicular triangle into the chest — is a fascial and bioelectrical propagation wave following the anatomical architecture of the deep cervical fascia and its continuity with the thoracic inlet fascia and anterior thoracic fascial layers.
The sequence of the cascade reflects the structural anatomy precisely:
The otic ganglion territory releases first. It is at the skull base juncture of the compressive forces — the most superior point held by the first hand, the point of maximum convergence of anterior and posterior crush forces. The piezoelectric current initiates at the deepest point of compression. The Ruffini mechanoreceptors at SJ 17 in the parotid and posterior infratemporal territory begin their sympathetic tone reduction. The suboccipital release at GB 12-20 begins reducing the posterior compressive force on the jugular foramen from above.
The cascade descends through the carotid sheath. Following the fascial sleeve that connects the skull base to the thoracic inlet — carrying the vagus nerve, carotid vessels, and cervical sympathetic trunk in a continuous fascial tube from the jugular foramen to the thorax. This is the primary anatomical channel of the descending cascade.
The cascade reaches the C4 thumb territory. The cardiac autonomic hub. HRV begins to shift — the carotid sinus sustained contact increasing vagal tone, the phrenic nerve decompression normalizing diaphragmatic motion and beginning to restore the respiratory sinus arrhythmia. The breathing changes as the diaphragm regains fuller motion. The cardiac rhythm begins to shift.
The cascade reaches the clavicular triangle. The second hand is receiving the descending wave and providing the inferior anchor that allows the release to complete rather than dissipate. The SCM shear at REN 22 provides the mechanical completion — the inferior attachment releasing as the superior attachment releases through the first hand’s contact, the entire muscle’s tension architecture releasing through the two-hand span simultaneously.
The cascade crosses the thoracic inlet. The clavicular contacts allow propagation from the cervical compartment into the thoracic compartment — into the anterior mediastinum and pericardial territory. Venous return begins to improve as diaphragmatic motion normalizes. Intrathoracic pressure dynamics shift. Vagal afferent input from diaphragmatic and pericardial mechanoreceptors begins to increase. The heart begins to receive more balanced autonomic input — more vagal, less sympathetic, better pericardial sensory feedback.
Getting It Out of the Head and Into the Chest
The cervical crush pattern traps the autonomic dysregulation above the thoracic inlet. The chronic jugular venous compression, the vagal compression, the carotid restriction, the scalene-thoracic outlet compression — all create a functional barrier at the thoracic inlet that prevents the normal craniocaudal flow of venous drainage, CSF pressure wave propagation, vagal signal propagation, and lymphatic drainage. The head becomes neurologically and hemodynamically congested above this barrier.
The two-hand technique simultaneously opens the superior end of the compressed cylinder with the first hand and the inferior end with the second — creating a pressure differential that allows the trapped contents to flow in the anatomically natural direction. The otic ganglion territory releasing is the superior valve opening. The clavicular triangle contacts are the inferior valve opening. The SCM shear at REN 22 is the final mechanical release of the anterior cervical fascial barrier.
The clinical observation — a secondary movement starting at the otic territory and descending through the clavicular triangle into the chest — is the direct sensory perception of this fascial and bioelectrical propagation wave following the deep cervical fascial architecture in its natural craniocaudal direction.
As the cascade reaches the pericardial territory, the Liver Jueyin not nourishing Heart pattern begins to resolve — not through energetic intervention in the abstract sense but through the specific mechanical, vascular, and autonomic pathways mapped in Article Two. The Heart begins to receive the vagal input that gives the pulse its full quality. The hidden pulse begins to emerge. The Yin-Yang integration function at the otic ganglion juncture begins to restore.
Part Nine: Practitioner State as Therapeutic Variable
The Practitioner’s Regulated Nervous System
Schore (2012) is explicit that therapeutic change for early relational trauma requires right-hemisphere-to-right-hemisphere transmission in a regulated relational field — the therapist’s own right hemisphere regulatory capacity as the instrument of change, not their technique.
In the context of this manual technique, this principle has a direct physiological expression through the HeartMath cardiac coherence mechanism described in Article Two. The practitioner’s cardiac coherence state during treatment is an electromagnetic input to the patient’s autonomic regulatory system operating through the cardiac field proximity transmission that McCraty et al. (2009) have documented. A practitioner in cardiac incoherence — anxious, rushed, focused primarily on correct finger placement rather than present-state regulation — is transmitting incoherence into the patient’s autonomic field simultaneously with the technique.
The technique operates through the tissue’s own piezoelectric and mechanoreceptor properties. The practitioner’s regulated nervous system operating through physical contact is amplifying or attenuating those tissue properties depending on its own coherence state. This is not metaphysical. It is the cardiac electromagnetic field transmission mechanism operating through the proximity of treatment contact.
Practical implication: the practitioner’s own coherence breathing practice — HeartMath protocol, or equivalent — is not a personal wellness activity separate from clinical competence. It is a clinical preparation activity directly relevant to treatment outcomes in this population.
Titration as Clinical Necessity
For the population described in Articles One and Two — with kindled PAG, retained primitive reflexes, encapsulated preverbal trauma, and hippocampal involution — titration is not a therapeutic nicety. It is a biological requirement.
The release initiated by this technique is opening access to material that the dissociative suppression system has been containing, sometimes for decades. The rate of release must be controlled such that:
The PAG does not cross its activation threshold — if the release proceeds faster than the regulatory capacity can contain, the system activates the freeze or flight response and shuts down the release.
The mPFC suppression does not overwhelm the emerging interoceptive signals — the system needs to be able to tolerate brief windows of increased body awareness as the cervical compression releases.
The person has enough regulatory capacity to stay with what arises rather than immediately re-dissociating.
The practical expression of titration in this technique is the five-second hold principle — allowing the tissue to pass through its elastic resistance phase into the viscoelastic melt and piezoelectric propagation phase, then following the tissue’s own rhythm rather than driving the release further. The tissue melt tells the practitioner the tissue is ready to move. The practitioner follows rather than leads.
Part Ten: Self-Care Adaptations
The Self-Application Principle
Several components of the two-hand technique are directly adaptable for self-care application. Given the population’s limited access to appropriate clinical care, the chronic nature of the presentation, and the frequency required to drive genuine autonomic plasticity — daily self-care practice addressing the same anatomical targets through adapted contact is clinically significant.
The Clavicular Triangle Self-Contact. The second hand position — middle finger along the clavicle, ring finger in the first intercostal space, featherweight contact at ST 12-13, pinky toward ST 14, thumb at REN 22 — is straightforwardly self-applicable. The ipsilateral hand can reach the clavicular triangle in a modified position. This contact applied for five minutes daily directly addresses the thoracic inlet fascial barrier, the stellate ganglion territory, the pectoralis minor compression, and the SCM sternal attachment. The five-second hold principle applies — sustained gentle contact, following the tissue’s response rather than forcing.
HeartMath Coherence Breathing. The 5-5 coherence breathing pattern — five-second inhalation, five-second exhalation — directly produces the respiratory sinus arrhythmia that increases HRV and builds vagal tone. Practiced for ten minutes daily, with attention placed at the cardiac region, it produces measurable HRV improvement within weeks (McCraty et al., 2009). It requires no contact, no equipment, no cortical processing of emotional content. It builds the parasympathetic baseline from which all other treatment benefits can compound.
The Diving Reflex. Cold water contact on the face — forehead, periorbital region, and nose — activates the TCR through the ophthalmic division of the trigeminal nerve, producing direct vagal cardiac activation through the oculocardiac reflex pathway. Cold water facial immersion or cold water splash to the forehead and periorbital region produces immediate HRV increase and vagal tone boost. Practiced as a morning reset — 20-30 seconds of cold water contact on the face — it provides a daily direct TCR-mediated parasympathetic activation accessible to anyone with running water.
Aerobic Exercise. The most robustly supported intervention for hippocampal neurogenesis — directly upregulating BDNF through aerobic activity with effect sizes documented within weeks of consistent practice (Erickson et al., 2011). For the population with hippocampal involution described in Article One, aerobic exercise is not general health advice. It is the most evidence-based specific intervention available for the hippocampal structural compromise that underlies the treatment resistance of this population. Thirty minutes of moderate aerobic activity five times weekly is the minimum effective dose in the neurogenesis literature.
Jaw and Suboccipital Self-Release. Gentle self-massage of the masseter, the temporal region (in the ST 7 territory), and the suboccipital muscles using sustained non-painful contact with the fingertips — following the tissue melt principle — addresses the two components of the cervical crush most accessible to self-treatment. Applying sustained contact for five to ten seconds and following the tissue softening, rather than rubbing or kneading, accesses the Ruffini mechanoreceptors rather than the nociceptive pathways. Performed before sleep it may also reduce bruxism-related V3 activation and improve the sleep architecture needed for hippocampal consolidation.
Sleep Architecture Support. The glymphatic system — which clears metabolic waste from the brain, including the waste products of the chronic neuroinflammatory load described in these articles — operates primarily during slow-wave sleep. The intracranial venous hypertension from jugular compression impairs CSF dynamics and glymphatic flow. As the cervical compression begins to release through clinical treatment and self-care, improving sleep depth and duration becomes a compounding benefit — each night of improved sleep contributing to the glymphatic clearance that reduces the neuroinflammatory load that was sustaining the sympathetic overdrive and the chronic cortisol elevation. Blue light elimination two hours before sleep, screen-free bedrooms, and consistent sleep timing to protect the melatonin-circadian axis are the minimum environmental conditions for this to occur.
Part Eleven: Contraindications and Clinical Flags
When to Modify or Refer
The anatomical territory addressed by this technique includes structures whose compromise requires clinical caution:
Carotid artery pathology. The carotid sinus contact at C4 is contraindicated in patients with known carotid stenosis, carotid plaque, or history of carotid intervention. Carotid sinus hypersensitivity — a condition producing exaggerated bradycardia or syncope with carotid sinus pressure — requires extreme caution or avoidance of the C4 thumb contact. Standard clinical screening for dizziness with cervical rotation and extension before treatment is appropriate.
Active inflammatory lesions in the craniofacial territory. Acute sinusitis, acute dental abscess, acute TMJ inflammation, or any active infectious process in the face or neck requires deferral until the acute phase resolves. Acute inflammation in the sphenoid or temporal regions in the context of known craniofacial trauma history requires medical clearance before work in those territories.
Acute cervical instability. Any cervical instability — atlantoaxial instability, fracture, acute disc herniation with radiculopathy — requires medical assessment before any cervical manual work. The gentle nature of the technique reduces but does not eliminate risk in this context.
Severe cardiac arrhythmia. The TCR activation potential of the otic ganglion territory contact requires caution in patients with known severe cardiac arrhythmia, complete heart block, or sick sinus syndrome. The technique produces vagal cardiac activation — which is therapeutic for the dysrhythmias of vagal insufficiency but potentially contraindicated in rhythm disturbances where vagal activation is itself destabilizing.
Thoracic outlet syndrome with vascular compromise. Known subclavian artery or vein compromise in the thoracic outlet requires modification of the clavicular contacts to avoid exacerbating vascular compression.
The PAG activation signs. During treatment, signs of PAG activation — sudden dissociation, color change (pallor or flushing), significant heart rate change, sudden temperature change, sudden somnolence, or sudden agitation — require immediate reduction of contact depth and a pause in technique. These signs indicate the release has exceeded the system’s current regulatory capacity. The titration principle requires working below the threshold of PAG activation consistently.
Conclusion: The Acupuncture Point Geography Was Always Telling Us
The face and skull acupuncture points sitting directly over the parasympathetic ganglia, trigeminal emergence points, cranial nerve exit foramina, and autonomic convergence points mapped across these three articles is not coincidental. Classical practitioners found these points through centuries of clinical observation because they were working with patients whose bodies responded to contact in these locations with consistent and powerful autonomic effects. The points found themselves.
What the current synthesis adds is the neuroanatomical explanation for why those points produce those effects — and the critical clinical implication that follows: the access method determines whether those effects are therapeutic or harmful in this specific population.
Touch accesses these points through the sensory pathways that do not trigger the threat detection cascade. Needles in this territory trigger that cascade through unavoidable nociceptive activation. Electrical stimulation compounds the harm through multiple converging mechanisms.
The self-care system emerging from this framework — coherence breathing, cold water diving reflex, aerobic exercise for hippocampal neurogenesis, sleep architecture support, and adapted self-contact of the clavicular triangle and jaw territory — provides a daily-dose intervention in the autonomic regulatory territory that clinical treatment opens but cannot alone sustain. The plasticity required to move these deeply established circuit organizations requires repetition on the scale of the original developmental experience. Clinical treatment provides the skilled opening. Self-care practice provides the daily repetition that drives the actual architectural change.
The population addressed by this framework — the three generations into inherited vagal depletion, the preverbal trauma survivors, the disorganized attachment adults navigating a world whose therapeutic systems have largely labeled their neurology as character — deserves both the clinical sophistication that meets them where they actually are, and the self-care tools that give them agency in their own restoration.
This series of articles is a preliminary mapping of that territory. It is offered as a clinical synthesis for practitioners working at the intersection of TCM, polyvagal neuroscience, developmental trauma, and somatic approaches — the practitioners who are already meeting this population and already observing the effects that the existing literature has not yet adequately explained.
References
Craig, A. D. (2003). Interoception: the sense of the physiological condition of the body. Current Opinion in Neurobiology, 13(4), 500–505.
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., Kim, J. S., Heo, S., Alves, H., White, S. M., Wojcicki, T. R., Mailey, E., Vieira, V. J., Martin, S. A., Pence, B. D., Woods, J. A., McAuley, E., & Kramer, A. F. (2011). Exercise training increases size of hippocampus and improves memory. Proceedings of the National Academy of Sciences, 108(7), 3017–3022.
Langevin, H. M. (2006). Connective tissue: a body-wide signaling network? Medical Hypotheses, 66(6), 1074–1077.
Levine, P. A. (2010). In an unspoken voice: How the body releases trauma and restores goodness. North Atlantic Books.
Lowe, R. (2016). Acupuncture point combinations: The key to clinical success (2nd ed.). Churchill Livingstone.
McCraty, R., Atkinson, M., Tomasino, D., & Bradley, R. T. (2009). The coherent heart: heart-brain interactions, psychophysiological coherence, and the emergence of system-wide order. Integral Review, 5(2), 10–115.
McGlone, F., Wessberg, J., & Olausson, H. (2014). Discriminative and affective touch: sensing and feeling. Neuron, 82(4), 737–755.
Myers, T. W. (2020). Anatomy trains: Myofascial meridians for manual and movement therapists. Churchill Livingstone.
Porges, S. W. (2011). The polyvagal theory: Neurophysiological foundations of emotions, attachment, communication, and self-regulation. Norton.
Schleip, R. (2003). Fascial plasticity: a new neurobiological explanation. Journal of Bodywork and Movement Therapies, 7(1), 11–19.
Schore, A. N. (2012). The science of the art of psychotherapy. Norton.
Thompson-Schill, S. L., Jonides, J., Marshuetz, C., Smith, E. E., D’Esposito, M., Kan, I. P., Knight, R. T., & Swick, D. (2002). Effects of frontal lobe damage on interference effects in working memory. Cognitive, Affective, and Behavioral Neuroscience, 2(2), 109–120.
van der Kolk, B. A. (2014). The body keeps the score: Brain, mind, and body in the healing of trauma. Viking.
