
Introduction: A Signaling System Trained Into Silence
Disgust is not a secondary emotion. It is not a social construction or a learned preference. It is a primary neurochemical circuit with a specific anatomy, a specific neurotransmitter profile, and a specific evolutionary function: to generate avoidance of stimuli that threaten the organism’s integrity. Understanding why that circuit fails in survivors of early relational trauma, and what the downstream consequences of that failure are across a lifetime of relationship-making, is the subject of this article.
The clinical picture is consistent enough across the developmental trauma population to constitute a recognizable pattern. Adults presenting with histories of chronic childhood interpersonal trauma, particularly where the primary threat was the caregiver rather than an external source, show a characteristic deficit in social discernment. They describe knowing intellectually that a relationship feels wrong while being unable to act on that knowledge. They report that the gut-level signal that should generate withdrawal simply does not fire, or fires too late, or fires and is immediately overridden by something that feels more compelling than the signal itself. When the neurobiology of disgust is mapped against the developmental conditions these patients grew up in, the mechanism of that deficit becomes precise.
The Neurochemistry of Disgust
Disgust activates a circuit distinct from the general threat-response circuitry. Its primary anatomical anchors are the left inferior frontal cortex, the left amygdala, and most critically the insular cortex, with the right putamen and left insula showing the most consistent activation across neuroimaging studies. Damage to the insula and putamen, as seen in Huntington’s disease, specifically impairs both the recognition and the experience of disgust while leaving other emotional responses relatively intact. This anatomical specificity is clinically important: disgust is not simply a variant of fear or aversion. It has its own dedicated hardware. (Sprengelmeyer et al., 1996; van der Kolk, 2014)
The dominant neurochemical modulator of that hardware is serotonin. High serotonin activity in the insular cortex regulates the intensity of disgust expression. The relationship is not simply that more serotonin produces more disgust. It is more precise than that: serotonin tone in the insula calibrates the threshold and intensity of the disgust signal, determining how readily the circuit fires and how strongly. Crockett et al., 2009; Porges, 2011) The inverse holds as well. Chronically low serotonin tone in the insular pathway produces a blunted or unreliable disgust signal, one that either fails to reach threshold or reaches it too late to function as genuine navigation.
How does that chronic deficit develop in the absence of any structural lesion? This is where the developmental picture becomes essential.
The Attachment Suppression Mechanism
A child whose primary caregiver is the source of chronic aversive input faces a neurobiological problem with no clean solution. The disgust circuit, which is functional from early development, generates a clear signal: this stimulus is harmful, withdraw, avoid. But the attachment system, which is equally fundamental and operates through entirely different circuitry, generates an opposing imperative: proximity to the caregiver is survival, maintaining the bond at all costs.
In adults encountering aversive stimuli from sources outside the attachment system, these two signals do not conflict. Disgust fires, avoidance follows, and the attachment system is not implicated. In a child whose attachment figure is simultaneously the source of the aversive input, the two systems are placed in direct opposition. The disgust circuit demands withdrawal. The attachment circuit demands proximity. The child cannot comply with both.
The resolution the developing nervous system arrives at is not a compromise. It is a suppression. The disgust signal is overridden, repeatedly, across thousands of interactions, throughout the developmental window when the insular-serotonergic pathway is still being calibrated. What begins as a situational suppression, required in this specific context with this specific person, becomes a trait-level baseline. The insular circuit learns through repeated developmental experience that its output will not be acted upon. Over time the signal degrades not because the circuit is structurally damaged but because the circuit has been trained into functional silence. (Schore, 2012)
The serotonin consequence of that training is direct. Chronic suppression of the insular disgust pathway dysregulates the serotonergic modulation of that system. The raphe nuclei, which are the primary serotonin-producing structures in the brainstem, calibrate their output in part through the feedback they receive from the systems they regulate. A system that is chronically suppressed provides a different feedback signal than a system that is allowed to function. The long-term result is a serotonin deficit in the insular pathway that is not incidental to the developmental trauma history. It is mechanistically produced by it. While the direct feedback loop between chronic insular suppression and raphe recalibration remains inferential, the well-documented role of serotonin in modulating aversive inhibition supports the broader neurochemical logic of this mechanism (Crockett et al., 2009).
This is not a purely psychological phenomenon. The same serotonin-insula pathway that generates gut-level social aversion also generates physical visceral revulsion. Many survivors of early relational trauma report that their physical gut signals are either absent or arrive far too late to be useful. The body-based signal that a regulated nervous system generates reliably in response to threat has been trained into unreliability. (van der Kolk, 2014) The practitioner who understands this mechanism stops attributing the patient’s repeated entry into harmful relationships to poor judgment or self-destructive motivation. The instrument of judgment has been compromised at the neurochemical level.
The Glycine-Raphe Interference: Disruption From Below
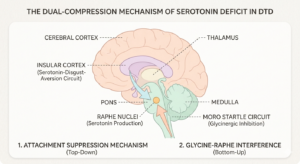 The serotonin deficit produced by the attachment suppression mechanism operates from above, through the repeated training of the insular pathway into silence. There is a second mechanism operating simultaneously from below, through the brainstem, that compounds the same deficit through a different route.
The serotonin deficit produced by the attachment suppression mechanism operates from above, through the repeated training of the insular pathway into silence. There is a second mechanism operating simultaneously from below, through the brainstem, that compounds the same deficit through a different route.
The startle reflex, specifically the Moro reflex in its unintegrated form in the DTD population, runs through the reticular formation and the cochlear nucleus. Its inhibitory modulation is heavily glycinergic. Glycine is the primary inhibitory neurotransmitter of the brainstem and spinal cord, functionally equivalent to what GABA provides at cortical levels. In a normally integrated startle response, glycinergic interneurons in the caudal brainstem damp the response after the initial firing. The circuit activates, the charge moves, and glycinergic damping brings the system back to baseline. (Lynch, 2004; Masgutova & Akhmatova, 2011)
When the Moro reflex remains unintegrated, as it does consistently in the DTD population, that glycinergic damping mechanism is chronically underperforming. The startle fires but does not complete its discharge cycle. The glycinergic interneurons that should close the loop are not doing their job, and the circuit remains in a state of incomplete activation.
Why does this matter for serotonin? Because the raphe nuclei, the primary serotonin-producing structures, sit within the reticular formation where the startle circuit runs. Glycine and serotonin interact directly at the level of the raphe. Chronic dysregulation of glycine cycling in the reticular formation chronically dysregulates raphe function. The serotonin system is being undermined at its source, not only through the top-down suppression of the insular pathway but through the bottom-up interference of the glycine-raphe interaction in a brainstem running an unintegrated startle.
The result is a system being compressed from two directions simultaneously. The attachment suppression mechanism trains the insular pathway into silence from above. The glycine-startle-raphe interference degrades the serotonin infrastructure from below. In a child with early relational trauma who also carries an unintegrated Moro, which is characteristic of the DTD population, both mechanisms are active together. The serotonin deficit is not incidental or mild. It is the product of two converging mechanisms that reinforce each other across the developmental window.
Downstream Consequences: Social Discernment and the Miscalibrated Picker
The clinical consequence that presents most consistently in the adult DTD population is not what it is most often named as. It is not poor judgment, low self-worth choosing what is familiar, or unconscious repetition of early relational patterns, though all of these may be present. The primary mechanism is neurochemical: the instrument that should generate this feels wrong is not generating reliably. The picker is miscalibrated because the serotonin-insula system that calibrates it has been operating at a deficit since early development.
This distinction matters clinically because it changes what the practitioner is working with. A patient whose social discernment failures are primarily a product of psychological pattern repetition is a different clinical problem from a patient whose serotonin-insula pathway has been in functional deficit since childhood. The first problem responds to insight and pattern interruption work. The second requires neurochemical substrate restoration before insight work has the infrastructure to land on. (Levine, 2010)
The same pathway that generates this person feels unsafe also generates the full range of embodied social attunement: the felt sense that someone is genuine, that an environment is safe, that a situation warrants trust. When the pathway is chronically blunted, what is lost is not just the negative signal. The positive signal is equally compromised. The result is the presentation that is so common in this population: a person who reports that relationships feel flat, that genuine connection seems either absent or arrives without the felt quality that should accompany it, that they know intellectually what closeness is supposed to feel like but cannot generate the embodied experience of it.
